

Irwin Stone era ingegnere chimico per formazione, biochimico per vocazione e paleopatologo per passatempo. Cominciò a lavorare sull’acido ascorbico nel 1934 e conseguì il primo brevetto USA per il suo uso come anti-ossidante alimentare. L’uso mondiale di questo processo è stato un fattore nella eliminazione virtuale dello scorbuto clinico conclamato nei paesi sviluppati.
Nel 1965-67 pubblicò una serie di articoli che descrivevano la malattia genetica, epato-enzimatica, Ipoascorbemia ed il suo significato in Medicina. Questo nuovo approccio genetico fornisce il razionale per l’uso di dosi massicce di acido ascorbico nella Terapia Ortomolecolare. E’ stato direttore della Megascorbic Research, Inc.
L’acido ascorbico è una sostanza ubiquitaria fondamentale nel processo vitale.
Tutti gli organismi viventi o la producono o la prendono dal nutrimento, altrimenti periscono. I sistemi enzimatici per la produzione dell’acido ascorbico sono di antica origine e si sono formati molto presto nello sviluppo del processo vitale su questo pianeta, probabilmente mentre le forme più altamente sviluppate erano ancora forme primitive unicellulari. L’evidenza della embriologia sia delle piante che degli animali corrobora questo punto di vista dal momento che il seme dormiente della pianta e l’uovo dell’animale sono privi di acido ascorbico. C’è una produzione immediata di acido ascorbico nel seme che germina o nell’uovo in sviluppo, perfino quando l’embrione non è niente più che un grumo di alcune cellule.
Anche la sua diffusa presenza in tutti gli odierni organismi pluricellulari, sia piante che animali, lo testimonia. Possiamo anche dedurre che la produzione di acido ascorbico era ben sviluppata prima che gli organismi viventi divergessero nelle forme di piante e di animali.
Stone.
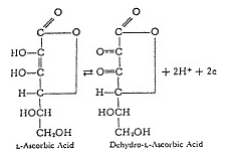
Dal punto di vista chimico, l’acido ascorbico è un semplice materiale carboidrato, relativo al glucosio, di proprietà alquanto uniche. La presenza del gruppo ene-diolo nella molecola conferisce la labilità degli elettroni, il che lo rende membro di un sistema ossido-riduttivo con proprietà di donazione-ricezione di elettroni, vedi fig. 1. A livello sub- molecolare, il processo vitale non è nulla più di un trasferimento di elettroni ordinato ed a passi, così che la presenza di un sistema elettrone- labile come l’acido ascorbico in un organismo vivente che agisce in concerto con altri antichi sistemi ossido-riduttivi, aiuta nel mantenimento della efficienza di trasferimento di elettroni nel processo vitale.
Se ci basiamo sull’esame degli animali oggi viventi per la loro produzione di acido ascorbico per stimare la sua presenza nei resti fossili, accumuliamo dati interessanti ed istruttivi sulla evoluzione dei sistemi enzimatici coinvolti. Sulla base della nota precisione del trasferimento genetico dell’informazione, possiamo assumere che i sistemi enzimatici e la loro localizzazione corporea negli odierni animali viventi è cambiata poco dai loro antichi rappresentanti.
Ciascuno dei primitivi invertebrati ed organismi inferiori esaminati finora ha mostrato la presenza di acido ascorbico, Bourne and Allen;2 Bourne.3 In quasi tutti i vertebrati esaminati la produzione di acido ascorbico è la norma. Quelle poche specie che non possono produrre il proprio acido ascorbico soffrono di un difetto genetico nei loro sistemi di produzione enzimatica e devono ricevere una fornitura di acido ascorbico dai loro cibi oppure muoiono di scorbuto.
La tavola 1 mostra la localizzazione del sistema ascorbico ed enzimatico nel corso della evoluzione dei vertebrati dai pesci ai primati, sulla base di dati ottenuti dall’esame delle odierne forme animali, Chatterjee et al.1 Roy and Guha5; Chaudhuri and Chatterjee.6. Il luogo degli enzimi per la produzione dell’acido ascorbico nei vertebrati a sangue freddo, i pesci, gli anfibi ed i rettili, è nei reni. I più altamente attivi mammiferi a sangue caldo tutti sintetizzano il loro acido ascorbico nel fegato. Una delle principali funzioni dell’acido ascorbico nella fisiologia animale è il mantenimento della omeostasi biochimica sotto stress. Più grande è lo stress a cui è sottoposto l’animale, più acido ascorbico produce.
Circa 165 milioni di anni fa, quando la natura aveva in vista l’evoluzione dei più attivi e stressanti mammiferi, doveva essere presa una importante decisione morfologica e fisiologica. I reni, pure adeguati come sito di sintesi dell’acido ascorbico per i lenti vertebrati a sangue freddo, erano inadeguati per le aumentate richieste di acido ascorbico per i più altamente stressati mammiferi. La soluzione vincente di questo problema fu il trasferimento degli enzimi per la produzione dell’acido ascorbico dai relativamente piccoli e biochimicamente affollati reni al più spazioso fegato, che è il più grande organo del corpo. Tutti gli odierni mammiferi capaci di sintetizzare acido ascorbico lo producono nel fegato perché ogni antica forma che non effettuò tale passaggio era così handicappata biochimicamente che fu eliminata dalle forze della evoluzione.
Gli uccelli odierni i cui antenati apparirono circa nello stesso tempo dei mammiferi, mostrano ancora questa transizione rene-fegato, Chaudhuri and Chatterjee. Il più vecchio ordine degli odierni uccelli, come le anatre, i piccioni ed i falchi, sintetizzano il loro acido ascorbico nei reni, mentre negli ordini più recenti degli uccelli da trespolo e canterini, i Passeriformi, alcuni producono acido ascorbico sia nei reni che nel fegato, altri solo nel loro fegato. Alcuni, come l’uomo, sono incapaci del tutto di sintetizzare acido ascorbico. Col procedere della evoluzione, i primati apparvero circa 65 milioni di anni fa e come gli altri mammiferi avrebbero dovuto essere capaci di sintetizzare l’acido ascorbico nel loro fegato. Comunque, qualche cosa accadde durante l’evoluzione dei primati perché è noto che da migliaia di anni l’uomo, a differenza degli altri mammiferi, era soggetto allo scorbuto.
Fino al 1907 lo scorbuto era considerato una malattia del tutto umana dal momento che nessun altro animale era noto essere suscettibile ad esso. Nel 1907, Holst and Frohlich , lavorando sullo beri-beri navale contratto a bordo della flotta da pesca norvegese, volevano un piccolo mammifero che sostituisse i piccioni allora usati come animali di prova. Essi nutrirono i porcellini d’India con la dieta di prova, che causava il beri-beri nei loro piccioni, e con loro grande sorpresa, ne risultò invece lo scorbuto. Più tardi fu mostrato che scimmie da laboratorio erano anche suscettibile allo scorbuto.
L’uomo, il porcellino d’India e certe scimmie, a differenza degli altri mammiferi, non possono produrre il loro acido ascorbico.
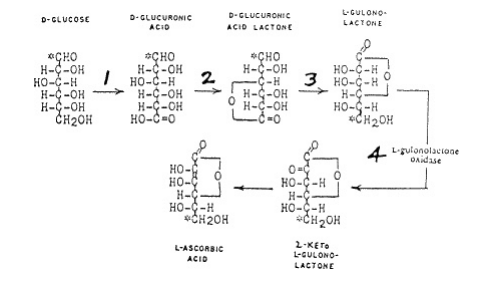
Nel 1912 fu postulata l’ipotesi delle vitamine (Funk), parte della quale stabiliva che lo scorbuto era una malattia da deficienza causata dalla mancanza nella dieta di una sostanza sconosciuta solubile in acqua, chiamata vitamina C. Venti anni più tardi nel 1932 (Svirbely and Szent-Györgyi ), fu dimostrato che l’acido ascorbico era identico alla vitamina C . Burns nel 1959 mostrò che la lesione biochimica basilare, nei pochi mammiferi soggetti allo scorbuto, era dovuta alla loro incapacità di produrre l’enzima attivo, L-gulonolactone ossidase, implicato nella conversione nei loro fegati del glucosio sanguigno dei mammiferi ad acido ascorbico. Questa sintesi, che implica quattro enzimi, è illustrata in fig. 2. L’uomo ha i primi tre enzimi nel suo fegato, ma gli manca il quarto enzima, il che blocca completamente la produzione di acido ascorbico da parte del fegato.
Fino al 1965 si assumeva che tutti i primati erano incapaci di produrre il proprio acido ascorbico ed erano soggetti alla malattia, lo scorbuto. Fu messo in evidenza (Stone), che questa era una mera assunzione che doveva essere provata. Fu suggerito che avrebbe dovuto essere esaminato l’intero ordine dei primati per la presenza dello Lgulonolactone ossidase nei loro fegati. Se questo fosse stato fatto, i dati ottenuti avrebbero potuto essere utili per indicare con esattezza, nel tempo, quando era intervenuta la mutazione. Così sarebbe stato possibile determinare in quale antenato primate dell’uomo si era perso questo importante enzima. Il suggerimento fu preso ed i test furono riportati da Harvard (Elliott et al.11) nel 1966 e dallo Yerkes Primate Research Center nel 1969 (Nakajima et al.12) dove si indicò che tutte le scimmie esaminate, che erano membri del sottordine, Prosimii, mostravano l’enzima L-gulonolactone ossidase attivo nei loro fegati, mentre nei fegati dei membri del sottordine Anthropoidea era inattivo. Mentre i dati non sono totalmente completi per tutto l’ordine dei primati, il presente campione indica che il tempo di separazione tra quei primati che sono soggetti allo scorbuto e quelli che non lo sono è probabilmente tra i due principali sottordini, Prosimii e Anthropoidea.
Ciò è illustrato in fig. 3 che mostra una carta dei resti fossili dei primati come concepito da Simons.13 La seconda colonna da destra è una lista degi odierni primati. La colonna all’estrema destra, marcata “Liver GLO” contiene i risultati dei recenti esami dei fegati dei primati per la presenza dello enzima attivo, L-gulonolactone ossidase. In tutti i primati esaminati finora, quelli nei sei generi del sottordine, Anthropoidea, mancano di questo enzima nei loro fegati mentre quelli dei quattro generi Prosimii hanno l’enzima attivo e possono sintetizzare il loro acido ascorbico. Sebbene altri membri delle famiglie Prosimii dovrebbero essere esaminati, come i Lemuridae, Tarsiidae e Daubentoniidae, le attuali indicazioni sono che la divisione tra i Prosimii e gli Anthropoidea non è solo morfologica ma anche in questo importante parametro fisiologico.
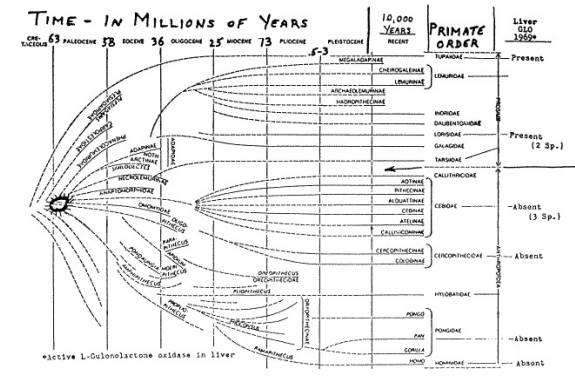
Se si segue la freccia e si estrapola questa linea divisoria all’indietro nel tempo in questa carta, allora si arriva al punto tra il tardo Cretaceo ed il tardo Paleocene dove questa mutazione, che distrusse il gene per la sintesi della proteina enzima Lgulonolactone ossidase, sembra sia avvenuta. Questo successe non molto dopo che i primati sono apparsi sulla scena, da 58 a 63 circa milioni di anni fa in vicinanza del cerchio stellato, nel primate antenato la cui progenie si è evoluta nel sottordine degli Anthropoidea.
E’ probabilmente più che una coincidenza che nelle vicinanze del periodo tardo Cretaceo, quando accadde questo incidente genetico nei primati, ci fu un breve intervallo in cui molti altri organismi incorsero in una severa attenuazione nella diversità o si estinsero.
Questo incluse molte forme di invertebrati e, da notare, i vertebrati dinosauri, che scomparvero improvvisamente dai resti fossili. Russell e Tucker nel 1971 suggerirono che l’esplosione di una supernova vicina, con la sua liberazione ed assorbimento da parte dell’atmosfera terrestre di grandi flussi di raggi gamma cosmici e di raggi x, potrebbero avere prodotto effetti climatici casì drastici da causare l’estinzione di molti animali inclusi i dinosauri, animali a sangue freddo. Se si verificò un tale evento astronomico, allora l’assorbimento casuale di parte di questa radiazione ad alto livello energetico potrebbe avere dato un contributo decisivo alla mutazione del gene dei primati per la sintesi della proteina enzima L-gulonolactone ossidase, col risultato di un enzima disattivo.
Pertanto una mutazione letale di condizione (Gluecksohn-Waelsch) accadde a questo primitivo primate. La distruzione di un tale processo biochimico vitale avrebbe avuto conseguenze letali se non fosse stato per il fatto che si presentò in un animale arboreo che viveva in un ambiente tropicale o semi- tropicale dove erano disponibili tutto l’anno molti cibi che contenevano acido ascorbico. La dieta del primate mutato forse non forniva altrettanto acido ascorbico quanto quello prima sintetizzato nel fegato, ma era sufficiente per sopravvivere. Bourne nel 1944 mostrò che un moderno gorilla, vivendo nel suo habitat naturale, otterrebbe 4.5 grammi di acido ascorbico al giorno dal suo cibo.
Pauling nel 1970, basando i suoi calcoli sul contenuto calorico e sui livelli di acido ascorbico nel cibi vegetali crudi, concluse che la gamma di assunzione ottima è circa da 2.3 a 9.5 grammi al giorno. Egli notò anche che, mentre la gamma delle vitamine B in 110 cibi vegetali crudi che forniscono 2500 Calorie era solo da due a quattro volte la dose giornaliera raccomandata nella dieta, il rapporto corrispondente per l’acido ascorbico era 35 volte quello raccomandato, 2300 milligrammi contro i 60 milligrammi al giorno.
Pauling nel 1968 indicò che questa mutazione può avere avuto valore di sopravvivenza a suo tempo perché liberò la macchina biochimica per altri scopi e conservava energia. Il valore di sopravvivenza fu perso non appena la progenie di questo animale mutato, evolvendo verso il futuro genere, Homo, lasciò gli alberi, si spostò verso climi temperati e cambiò la sua dieta in una in cui alti livelli di acido ascorbico non dominavano per tutto l’anno.
L’uomo porta ancora questo gene difettoso ed esso non ha valore di sopravvivenza per l’Uomo moderno. Infatti, nel corso della preistoria e durante i tempi storici, ciò ha costituito un handicap severo e gli effetti collaterali di questo gene difettoso sono risultati nella morte di più individui, nella causa di più malattie e sofferenze ed ha cambiato il corso della storia più di ogni altro singolo fattore.
Come risultato di questi studi evolutivi, Stone nel 1966 indicò che a causa di questo gene difettoso, che produceva un enzima inattivo, gli odierni umani soffrono di un innato errore dei mammiferi nel metabolismo dei carboidrati che fu chiamato Ipoascorbemia. Lo scorbuto è di fatto la conseguenza di questa malattia genetica epato-enzimatica che rende per l’uomo necessario ottenere acido ascorbico da sorgenti esterne. Negli ultimi 60 anni, l’acido ascorbico è stato visto come “vitamina C” quando in realtà è un metabolita del fegato e certamente non una “vitamina” per la miriade di mammiferi che hanno intatto il gene per lo Lgulonolactone ossidase. Tali mammiferi non prendono lo scorbuto, anche se hanno una dieta completamente priva di vitamina C. Se si definisce la correzione di questo innato errore nel metabolismo dei carboidrati come il bisogno di fornire, all’individuo, l’ammontare giornaliero di acido ascorbico che il fegato umano produrrebbe, se fosse presente il gene intatto, allora la produzione giornaliera di questo metabolita da parte del fegato sarebbe da due a quattro grammi, sulla base dei dati del ratto non stressato (Bur ns et al.21 Saloman and Stubbs) e almeno di 15 grammi al giorno sotto stress, Conney et al.23
Se si compara questa sintesi giornaliera del “metabolita del fegato”, acido ascorbico, con la dose giornaliera di “vitamina C”, 60 milligrammi (Food and Nutrition Board), raccomandati come nutrizionalmente adeguati per gli umani, c’è una disparità da 33 a 250 volte. Fin da quando fu postulata nel 1912 l’ipotesi di questo errore dietetico- vitamina C, l’enfasi fu posta sulla prevenzione o sulla cura dei sintomi dello scorbuto conclamato, così trascurando le molte altre funzioni basilari dell’acido ascorbico nel processo vitale. Un soggetto a cui si dà giornalmente la dose di pochi milligrammi di vitamina C non mostrerà i classici sintomi dello scorbuto clinico conclamato ma potrebbe tuttavia soffrire di un severo scorbuto sub-clinico. L’ipotesi vitamina C, che ebbe incontestata accettazione fin dal 1912, è stata originata in un tempo in cui i moderni concetti biochimici e genetici largamente accettati, erano o sconosciuti o non riconosciuti. Le classiche lezioni di Sir Archibald Garrod sugli “Errori Innati del Metabolismo,” in cui egli mostrava che la mancanza di enzimi poteva causare malattie, si tennero solo quattro anni prima, nel 1908. Questo nuovo e rivoluzionario concetto medico fu ignorato e trascurato per decenni. La scoperta, l’isolamento e la sintesi dell’acido ascorbico sarebbero giunte tuttavia venti anni dopo.
Le moderne concezioni del trasferimento della informazione genetica erano sconosciute. Chiaramente l’ipotesi vitamina C vecchia di 60 anni con il suo sottinteso orientamento verso dosi minuscole e l’allontanamento dello scorbuto clinico acuto è una grossolana semplificazione della nostra attuale conoscenza e subordina gli effetti a lungo termine delle croniche inadeguate assunzioni di acido ascorbico alla moltitudine di funzioni fisiologiche dell’acido ascorbico nel corpo umano.
Nella ricerca clinica degli ultimi 40 anni ci sono stati centinaia di articoli scritti per cercare il livello minimo giornaliero di acido ascorbico necessario per prevenire o eliminare i classici sintomi dello scorbuto clinico conclamato. Comunque, non trovereste un singolo articolo che riporti gli effetti a lungo termine di una continua somministrazione di acido ascorbico basata sui livelli prodotti nel tempo internamente nel fegato dei mammiferi. Questi alti livelli di produzione di acido ascorbico sono serviti molto bene nel mantenere la omeostasi biochimica sotto stress negli ultimi 165 milioni di anni durante l’evoluzione ed il dominio mondiale dei mammiferi. Dopo circa 60 milioni di anni dall’avvento della mutazione nei primati, è solo negli ultimi 40 anni, dalla produzione sintetica di illimitate quantità di acido ascorbico, che è diventato possibile correggere completamente lentamente questa malattia genetica enzimoepatica.
Questi concetti genetici forniscono il razionale per l’uso di dosi massicce (livelli di megascorbato) di acido ascorbico e aprono grandi orizzonti di ricerca nella medicina preventiva e nella terapia. Nelle poche aree in cui la terapia megascorbica è stata provata, è stata eminentemente un successo: nel trattamento delle malattie virali, nel glaucoma, nella schizofrenia come disintossicante. La ricerca su altre applicazioni di questi principi gene tici porterà alla profilassi megascorbica ed alla terapia megascorbica di molti altri stati di malattia.
1.1.1 Sommario e Conclusioni
Si è presentata la prova che i sistemi enzimatici funzionanti nel tempo per la importante sintesi dell’acido ascorbico sono di origine molto antica, cominciando molto prima del tempo in cui le linee delle piante e degli animali divergessero. La evoluzione progressiva di questi enzimi nei vertebrati è tracciata attraverso i pesci, gli anfibi, i rettili, gli uccelli ed i mammiferi. Poco dopo l’apparizione dei primati, ci fu una mutazione genetica del gene per l’enzima epatico L-gulonolactone ossidase, che distrusse la capacità animale di produrre acido ascorbico a partire dal glucosio del sangue. La progenie di questo animale mutato si sviluppò nei membri odierni del sottoordine dei primati Anthropoidea. I primati non mutati sono gli antenati dell’odierno ordine dei Prosimii.
L’estrapolazione di questi dati nei resti fossili dei primati indica che la mutazione è avvenuta nello stesso periodo della esplosione di una supernova vicina che forse fu responsabile dell’estinzione dei dinosauri e della scomparsa di molti invertebrati nel tardo Cretaceo. I membri del genere Homo, l’Uomo odierno, presentano ancora questo gene difettoso e durante i tempi preistorici e storici esso è stato responsabile di molte morti, molte malattie e sofferenza e di molti cambiamenti storici, più di ogni altro singolo fattore. L’importanza vitale dell’acido ascorbico in molte fasi della fisiologia umana è stata sottostimata negli ultimi 60 anni perché nel 1912, 20 anni prima della sua scoperta e sintesi, fu designata come “vitamina” per il trattamento dello scorbuto clinico conclamato, che era considerato un semplice disturbo dietetico.
In realtà l’acido ascorbico è un metabolita epatico prodotto in quasi tutti i mammiferi in grandi quantità giornaliere. A causa di questo gene difettoso l’Uomo soffre di una malattia epato-enzimatica genetica nei mammiferi, un vero “errore innato nel metabolismo dei carboidrati”, detto Ipoascorbemia. Lo scorbuto non è una distinta entità di malattia, ma meramente la sequenza finale fatale della Ipoascorbemia non trattata. L’approccio genetico fornisce il razionale per l’uso di grandi dosi giornaliere di acido ascorbico ed apre larghe prospettive di ricerca per la sua applicazione alla medicina preventiva ed alla terapia.
RIFERIMENTI
1. STONE, I.: Studies of a mammalian enzyme system for producing evolutionary evidence in Man. Amer. J. Phys. Anthrop. 3:83-85, 1965,
2. BOURNE, G. and ALLEN, R.: Vitamin C in lower organisms. Nature, 136:185-186, 1935.
3. BOURNE, G.: Vitamin C in the Australian Fauna. Med. J. Aust. 3:260-261, 1935.
4. CHATTERJEE, I. B., KAR, N. C., GHOSH, N. C. and GUHA, B. C.: Aspects of ascorbic acid biosynthesis in animals. Ann. N.Y. Acad. Sc. 92, Art. 1:36-56, 1961.
5. Roy, R. N. and Guha, B. C.: Species difference in regard to the biosynthesis of ascorbic acid. Nature, 182:319-320, 1958.
6. CHADHURI, C. R. and CHATTERJEE, I. B.: Ascorbic acid synthesis in birds: Phylogenetic trend. Science, 164:435-436, 1969. [*]7. HOLST, A. and FROHLICH, T.: Experimental studies relating to ship beriberi and scurvy, II. On the etiology of scurvy. J. Hyg. 7:634-671, 1907.
8. FUNK, C.: The etiology of the deficiency diseases. J. State Med. 20:341-368, 1912.
9. SVIRBELY, J. V. and SZENT-GYÖRGYI, A.: Hexuronic acid as antiscorbutic factor Nature, 129:690, 1932.
10. BURNS, J. J.: Biosynthesis of L-ascorbic acid; basic defect in scurvy. Amer. J. Med. 26:740-748, 1959.
11. ELIOTT, O., YASS, N. J. and HEGSTED, D. M.: Biosynthesis of ascorbic acid in the tree shrew and slow loris. Nature 212:739-740, 1966.
12. NAKAJIMA, Y., SHAUTHA, T. R. and BOURNE, C. H.: Histochemical detection of L-gulonolactone. Histochemie 18:293-301, 1969.
13. SIMONS, E. L.: The early relatives of man. Sc. Amer. pp. 55, July 1964.
14. RUSSELL, D. and TUCKER, V. V.: Supernovae and the extinction of the dinosaurs. Nature 229:553-554, 1971.
15. GLUECKSOHN-WAELSCH: Lethal genes and analysis of differentiation. Science 142:1269-1276, 1963.
16. BOURNE, C.: Vitamin C and immunity. Brit. J. Nutr. 2:341, 1949.
17. PAULING, L.: Evolution and the need for ascorbic acid. Proc. Nat. Acad. Sc. 67:1643-1648, 1970.
18. PAULING, L.: Orthomolecular Psychiatry. Science, 160:265-271, 1968.
19. STONE, I.: On the genetic etiology of scurvy. Acta Genet. Med. et Gemel. 15:345-350, 1966.
20. STONE, I.: Hypoascorbemia, the genetic disease causing the human requirement for exogenous ascorbic acid. Pers. Biol. Med. [*]10:133-134, 1966.
21. BURNS, J. J., MOSBACH, E. H. and SCHULENBERG, S.: Ascorbic acid synthesis in normal and drug-treated rats studied with ascorbic-1-C14 acid. J. Biol. Chem. 207:679-687, 1954.
22. SALOMAN, L. I. and STUBBS, D. W.: Some aspects of the metabolism of ascorbic acid in rats. Ann. N.Y. Acad. Sc. 92: Art. 1:128-140, 1961.
23. CONNEY, A. H., BRAY, C. A., EVANS, C. and BURNS, J. J.: Metabolic interactions between l-ascorbic acid and drugs. Ann. N.Y. Acad. Sc. 92; Art. 1:115-127, 1961.
24. Food and Nutrition Board, National Research Council: Recommended daily allowances. Publication No. 1694. Nat. Acad. of Sciences, Washington, D.C., 1968.
25. GARROD, A. E.: Inborn errors of metabolism. Lancet 2:1-7, 73-79, 142-148. 214-217, 1908.
26. STONE, I.: The genetic disease, Hypoascorbemia. A fresh approach to an ancient disease and some of its medical implications. Acta Genet. Med. et Gemel. 16:52-62, 1967.
27. STONE, I.: The Healing Factor. “Vitamin C” Against Disease. Grosset & Dunlap, Inc., New York, 1972.
Da Orthomolecular Psychiatry, 1972, Volume 1, Numbers 2 & 3, pp. 82-89.